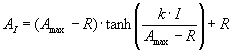
Individuenorientierte Prozessmodellierung (8 Stunden)
Betreuung: C. Eschenbach (Universität Hohenheim, Institut für Botanik) und W. Kurth
Einordnung in die Lehrveranstaltung: Die neue Forschungsrichtung der "Struktur-Funktions-Modelle" von Pflanzen, die sich in jüngster Zeit mehr und mehr international etabliert (Kongresse in Helsinki, Clermont-Ferrand und Montréal), erfordert die Verknüpfung von physiologischen Prozessinformationen mit Daten zur Pflanzenstruktur und ihrer Dynamik. In der Lehrveranstaltung sollen, über die reine Strukturmodellierung mit Lindenmayer-Systemen hinausgehend, auch hierzu sowie zur Frage des "upscaling" von Einzelorganen auf Individuen und Bestände Grundkenntnisse vermittelt werden.
Aktueller Hintergrund: Eine weitere Motivation für die Problemstellung des "upscaling" von Prozessinformationen und Stoffflüssen ergibt sich aus der Diskussion zur Klimaänderung. Der anthropogen bedingt erhöhte CO2-Gehalt in der Atmosphäre führt zu einer globalen Erwärmung (Treibhauseffekt, "global climatic change"). Viele Menschen sind in der Folge z.B. durch Meeresspiegelanstieg und Überschwemmungen bedroht. Auf internationalen Klimakonferenzen wird auf Abhilfe gesonnen und neben der Verpflichtung zur Reduzierung des CO2-Ausstoßes auch eine Wiederaufforstung diskutiert, um "überschüssiges" CO2 durch die Photosynthese der Bäume zu binden. Dazu ist es notwending, die CO2-Bindungskapazität der Wälder zu kennen. Prognosen zur großräumigen und langzeitigen Entwicklung der CO2-Bindung werden aus der Anwendung von Simulationsmodellen auf der Grundlage von Einzelmessungen abgeleitet. Hierzu ist das "upscaling" von einzelnen Pflanzenorganen zu individuellen Pflanzen und schließlich zu Beständen essentiell.
Übungssziel: Durch die zwei Übungstage in Stuttgart-Hohenheim soll ein kleiner (und naturgemäß oberflächlicher) Einblick in die Methoden der prozessorientierten Modellierung von Stoffflüssen vermittelt werden, welche zugleich einen wichtigen Teilaspekt von Struktur-Funktions-Modellen (z.B. ALMIS (Eschenbach 2000), Grocom (Kurth 1999)) darstellen. In einem ersten Schritt werden aus Messdaten Abhängigkeiten analysiert und mathematisch beschrieben. Mit Hilfe des so gewonnenen einfachen Modells in einer zweiten Übungsphase einfache Simulationsrechnungen durchgeführt werden. Als Beispiel dient der Gasaustausch eines Erlenbaums mit der Atmosphäre. Durch einen Vergleich mit der Atmung des Menschen einerseits und den Abgasen von Autos andererseits ergibt sich ein Gefühl für die Größenordungen, mit der jeweils ein Austausch mit der Atmosphäre stattfindet.
Zur Methode der bottom up-Modellierung: Der pflanzliche Gaswechsel hängt von endogenen (Entwicklungszustand, Adaptationstyp, diurnale Rhythmik) und exogenen Faktoren (mikroklimatische Faktoren in der Blattumgebung, Wasserhaushalt der Pflanze, u.ä.) ab. Um das Multifaktorensystem der Abhängigkeiten Schritt für Schritt zu erfassen, wird jeweils der quantitative Zusammenhang zwischen der Größe (Dosis) nur eines Faktors und seiner physiologischen Wirkung vor dem Hintergrund der Wirkung der anderen konstant gehaltenen Einflussfaktoren bestimmt. Für diese Abhängigkeit wird eine mathematische Gleichung angepasst. Auf der Grundlage dieser mathematischen Funktionen zur Beschreibungen der Abhängigkeiten des CO2-Gaswechsels von den Einflussfaktoren erfolgt die Modellierung der Kohlenstoffaufnahme der Blätter. Für die Hochrechnung von der Ebene der Einzelbätter auf die Ebene der ganzen Pflanze oder eines Pflanzenbestand sind zudem Daten zur Belaubung und über das unterschiedliche Verhalten der Blätter in den verschiedenen Bereichen eines Bestandes (z.B. Sonnen-, Schattenblätter) notwendig. Diese Informationen können in einem Struktur-Funktions-Modell des Pflanzenindividuums zusammengefasst werden. Die Behandlung von Struktur- und Prozessinformationen im Modell ALMIS wird übersichtsartig vorgestellt.
Literaturauswahl:
BEYSCHLAG, W., LANGE, O.L. & TENHUNEN, J.D. (1986): Photosynthese und Wasserhaushalt der immergrünen mediterranen Hartlaubpflanze Arbutus unedo L. im Jahreslauf am Freilandstandort in Portugal. I. Tagesverläufe von CO2-Gaswechsel und Transpiration unter natürlichen Bedingungen. Flora 178, 409-444 .
CALDWELL, M.M., MEISTER, H.-P., TENHUNEN, J.D. & LANGE, O.L. (1986): Canopy structure, light microclimate and leaf gas exchange of Quercus coccifera L. in a Portuguese macchia: measurements in different canopy layers and simulations with a canopy model. Trees 1, 25-41.
SCHULZE, E.-D. (1970): Der CO2-Gaswechsel der Buche (Fagus silvatica L.) in Abhängigkeit von den Klimafaktoren im Freiland. Flora 159, 177-232 .
Eschenbach, C. (1996): Zur Modellierung der Primärproduktion der Schwarzerle (Alnus glutinosa). Ecosys (Kiel).
ESCHENBACH, C. (2000): The effect of light acclimation of single leaves on whole tree growth and competition - an application of the tree growth model ALMIS. Ann. For. Sci. 57, 599-609.
KURTH, W. (1999): Die Simulation der Baumarchitektur mit Wachstumsgrammatiken. Wiss. Verlag Berlin; hier insbes. S. 238-242.
Teil 1
In der Übung wird nur der Einfluss der Einstrahlung und der Temperatur auf die Nettophotosynthese der Blätter analysiert. Ziel ist es, die unten angegebenen Funktionen für die Abhängigkeit der Nettophotosynthese von diesen beiden Faktoren an die Daten anzupassen und das Ergebnis graphisch darzustellen. Wir erhalten ein einfaches Simulationsmodell.
Die Einzelschritte:
1. Datengrundlage für die Analyse sind an Erlenblättern gemessene Tagesgänge des CO2-Gaswechsels und der Einstrahlung und Temperatur in der Blattumgebung. Die Datensätze wurden einerseits in der sonnenexponierten Kronenperipherie und andererseits im beschatteten Kroneninneren gemessen.
2. Um Quereffekte von Temperatur und Einstrahlung bei der Analyse der Abhängigkeiten zu vermeiden, müssen die Daten zunächst nach Temperatur- bzw. Lichtklassen sortiert werden.
3. Die Lichtsättigungsfunktion (Gleichung 1) wird mit der "curve fit -Funktion" des Programms Sigmaplot an die Daten in verschiedenen Temperaturbereichen angepasst. Die Werte für Amax, k und Rd sind zu bestimmen. Die gefundenen Kurven werden gemeinsam mit den Messwerten graphisch dargestellt.
4. Die Optimumfunktion für die Abhängigkeit der Nettophotosynthese von der Temperatur (Gleichung 2) wird mit der "curve fit -Funktion" des Programms Sigmaplot an die Daten in verschiedenen Lichtbereichen angepasst. Die Werte für Topt und Tmin sind zu bestimmen. Die gefundenen Kurven werden gemeinsam mit den Messwerten graphisch dargestellt.
5. Die durch "curve-fitting" gefundenen Abhängigkeiten der Nettophotosynthese der Blätter von der Einstrahlung und der Temperatur werden in einem Modell kombiniert.
6. In einem 3D-Plot kann die Abhängigkeiten der Nettophotosynthese gleichzeitig von der Einstrahlung und von der Temperatur dargestellt werden. Das "Käfig"-Diagramm zeigt die Nettophotosynthese-Werte für alle potentiell möglichen Kombinationen von Einstrahlung und Temperatur.
Gleichung (1):
|
AI |
lichtabhängige Nettophotosyntheserate bei gegebener Temperatur (µmol CO2 m-2 s-1) |
|
Amax |
maximale Nettophotosyntheserate (µmol CO2 m-2 s-1) |
|
R |
Dunkelatmung bei gegebener Temperatur (µmol CO2 m-2 s-1) |
|
k |
"Quantenausbeute", Anfangssteigung (µmol CO2 µmol Photonen-1) |
|
I |
Einstrahlung (µmol Photonen m-2 s-1) |
Gleichung (2):
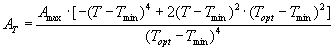 :
:
|
AT |
temperaturabhängige Nettophotosyntheserate bei sätt. Einstrahlung (µmol CO2 m-2 s-1) |
|
A max |
Nettophotosyntheserate im Scheitelpunkt der Kurve (µmol CO2 m-2 s-1) |
|
Topt |
Optimum-Temperatur (im Scheitelpunkt der Kurve; °C) |
|
Tmin |
unterer Temperaturkompensationspunkt (°C) |
|
T |
Temperatur (°C) |
Teil 2
Mit Hilfe des in Teil 1 erstellten Modells soll anhand von gemessenen mikroklimatischen Datensätzen und Belaubungsdaten der Gaswechsel eines Erlenbaums errechnet werden. Vergleichen Sie anschließend die Größenordnungen des menschlichen Atemumsatzes und der CO2-Abgase eines Mittelklassewagens mit den erechneten Werten. (Aber Vorsicht: In diesem sehr einfachen Modell werden viele andere Einflussfaktoren, die die Photosynthese reduzieren, sowie die Atmung von Stamm und Ästen nicht berücksichtigt. Die CO2-Fixierung des Baumes wird daher überschätzt, es handelt sich nicht um "die Wahrheit", sondern nur um Aussagen über die Größenordnung der Werte!)
Grundlagen für die Berechnung:
1. Datengrundlage für die Modellierung sind in verschiedenen Höhen in einem Erlenwald gemessene Tagesgänge der Einstrahlung und der Temperatur. Die Datensätze wurden einerseits in der sonnenexponierten Kronenperipherie (17 m Höhe) und andererseits im beschatteten Kroneninneren (11 m Höhe) gemessen.
Für eine ausgewachsene Erle im Bestand, die 18 m hoch und 52 Jahre alt war, wurde eine Gesamtblattfläche von 97 m2 bestimmt. In einem vereinfachten Ansatz ist ein Drittel der Blattfläche der Kronenperipherie zuzurechnen, und die restlichen Blätter sollen die Einstrahlung des beschatteten Kroneninneren erhalten.
2. Ein erwachsener Mensch hat in der Ruhe einen Sauerstoffverbrauch von etwa 0,3 l kg-1 Std -1 (bei sehr anstrengender Arbeit kann dieser bis zum zehnfachen Wert ansteigen), entsprechend 0,35 l min-1 für einen Erwachsenen von durchschnittlichem Gewicht (70 kg).
Das Verhältnis der Menge (Volumen) des ausgeatmeten Kohlendioxids zur Menge (Volumen) des aufgenommenen Sauerstoffs (= der respiratorische Quotient) liegt beim normal ernährten Menschen bei etwa 0,9.
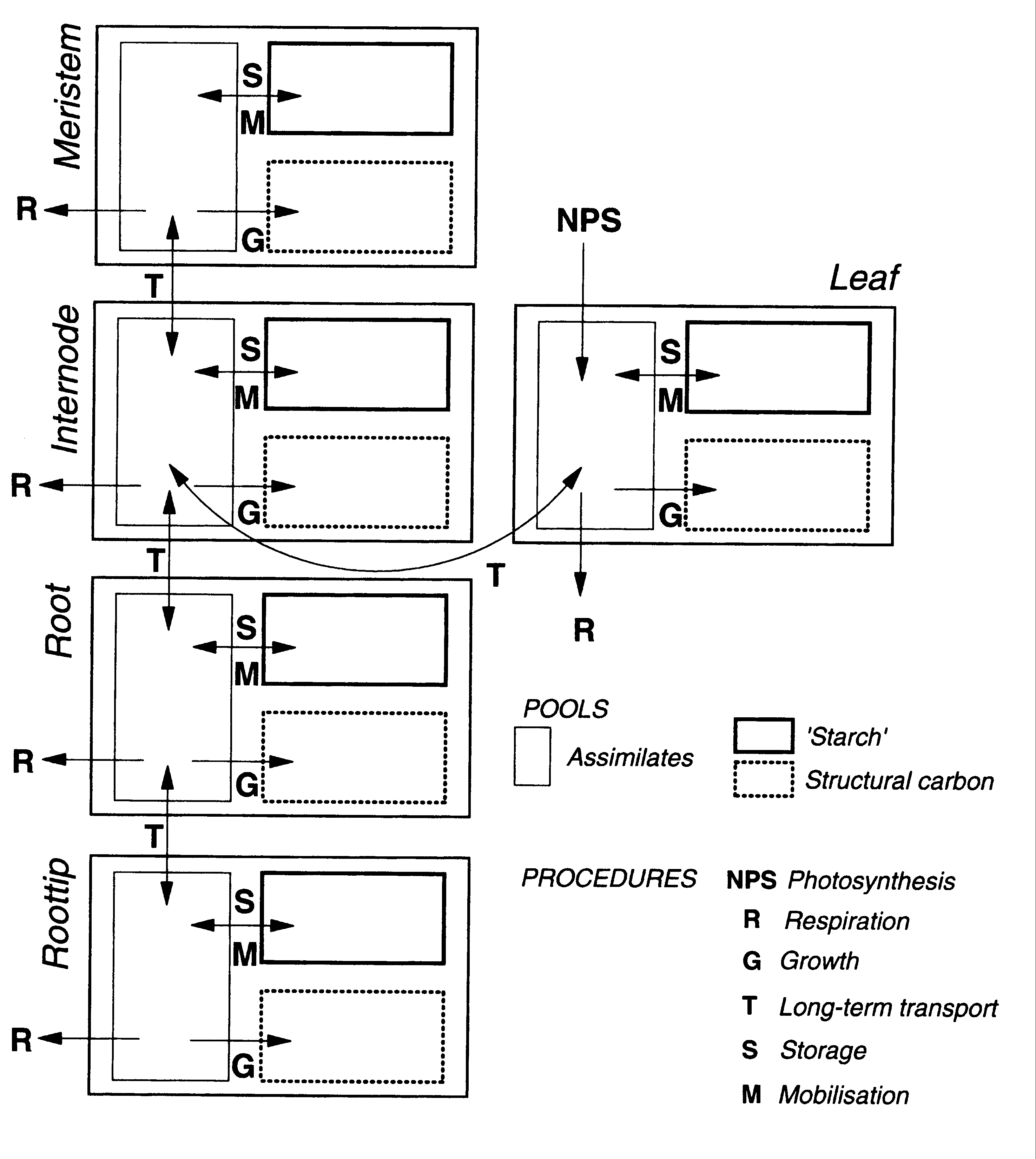
Teil 3Es wird die Repräsentation des organbezogenen Gaswechsels und der Pflanzenarchitektur-Informationen im Struktur-Funktions-Modell ALMIS erläutert und anhand von Modell-Läufen demonstriert. Grundlagen der objektorientierten Modellierung werden vorgestellt und am Beispiel des Gaswechselmodells erläutert. Das upscaling von den Pflanzenorganen zum Individuum und zur Baumgruppe wird in Simulationsläufen demonstriert.
Verknüpfung von Objekten und Prozessen im Modell ALMIS:
Last modification: May 16, 2001